- Home
- About Journals
-
Information for Authors/ReviewersEditorial Policies
Publication Fee
Publication Cycle - Process Flowchart
Online Manuscript Submission and Tracking System
Publishing Ethics and Rectitude
Authorship
Author Benefits
Reviewer Guidelines
Guest Editor Guidelines
Peer Review Workflow
Quick Track Option
Copyediting Services
Bentham Open Membership
Bentham Open Advisory Board
Archiving Policies
Fabricating and Stating False Information
Post Publication Discussions and Corrections
Editorial Management
Advertise With Us
Funding Agencies
Rate List
Kudos
General FAQs
Special Fee Waivers and Discounts
- Contact
- Help
- About Us
- Search


The Open Cell Signaling Journal
(Discontinued)
ISSN: 1876-3901 ― Volume 4, 2012
Pleiotropic Effect of Aldosterone on Human Endothelial Cells In Vitro
Pezhman Mirshahi, Alexandre Berthaut, Elodie Ducros, Jeannette Soria, Manjul K Agarwal, Massoud Mirshahi*
Abstract
Aim:
To investigate the effect of aldosterone and its antagonists on cell behavior and gene modulation in human endothelial cells.
Methods and Results:
Exposure of HBMEC to 100 nM aldosterone reduced the subsequent organization and assembly of cells into capillary-like networks, capillary length and cell multiplication but cell migration to the wound edge was not affected by the agonist. Eplerenone (400 nM) partially reversed the inhibitory effect of the agonist on capillary length, the number of capillary networks, as well as cell multiplication; the antagonist also inhibited the migration of HBMEC in a wound healing assay. Aldactone was more potent than eplerenone in most of the tests, due possibly to the fact that the former derivative is not specific to the MCR but activates other classes of steroid receptors as well. Transcriptional modulation by aldosterone was analyzed using a gene array technique that screened 1800 genes related to cytokines, monokines, growth factors, angiogenic effectors, cell metabolism, growth and malignant transformation. Evidence is provided here for the simultaneous upregulation of 36 mRNAs, and concurrent downregulation of 29 mRNAs, in endothelial cells exposed for 8 h to 100 nM aldosterone.
Conclusions:
Aldosterone inhibits cell migration, network formation, and cell proliferation in vitro, possibly via a pleiotropic effect on transcription modulation in human endothelial cells.
Article Information
Identifiers and Pagination:
Year: 2012Volume: 4
First Page: 1
Last Page: 8
Publisher Id: TOCELLSJ-4-1
DOI: 10.2174/1876390101204010001
Article History:
Received Date: 18/11/2011Revision Received Date: 2/12/2011
Acceptance Date: 16/12/2011
Electronic publication date: 30/1/2012
Collection year: 2012
open-access license: This is an open access article licensed under the terms of the Creative Commons Attribution Non-Commercial License (http: //creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted, non-commercial use, distribution and reproduction in any medium, provided the work is properly cited.
* Address correspondence to this author at the UMRS 872, Centre de Recherches des Cordeliers, Faculté de Médecine Paris VI, 15 rue de l’Ecole de Médecine, 75006 Paris, France; Tel: 00 33 1 42 34 69 31; Fax: 00 33 1 44 27 81 61; E-mails: dizbad2004@yahoo.fr, massoud.mirshahi@upmc.fr
| Open Peer Review Details | |||
|---|---|---|---|
| Manuscript submitted on 18-11-2011 |
Original Manuscript | Pleiotropic Effect of Aldosterone on Human Endothelial Cells In Vitro | |
INTRODUCTION
In the past, the body vasculature had generally been written off as a system of tubules connected end to end merely to facilitate blood flow. Of late, it has been recognized that 900 m2 or so of blood vessels form an important organ which assures body integrity in a number of ways [1-3]. Increasing evidence suggests that dysregulation of vessel growth underlies a myriad of disorders [3-5]. Inhibition of angiogenesis or capillary formation by angiostatin and endostatin is highly effective in tumor regression. Rheumatoid arthritis, diabetic retinopathy and psoriasis can also benefit from a similar therapy [6, 7]. On the other hand, angiogenic stimulation may be useful in ischemic diseases and decubitus [8, 9]. It therefore becomes important to delineate those factors that assure the genesis, maintenance, and renewal of body vasculature in health and disease.
The main, biologically active, component in the blood vessels consists of a single layer of endothelial cells that are insulated by an outer envelope consisting of pericytes, smooth muscle fibers, fibroblasts, the basement membrane, and an extracellular matrix [1]. Vascular endothelium is intimately linked to capillary formation, repair and remodeling of microcirculation [2]. It is well established that adrenocortical hormones exert pleiotropic effects on a wide variety of cell types [10, 11] and influence various parameters in the cardiovascular tree [12-14]. We had previously shown [16, 17] that the mineralocorticoid receptor (MCR)-mediated action of steroid hormones induces neosynthesis of the epithelial sodium channel [15] and also increases the cell volume of capillaries formed by human bone marrow endothelial cells (HBMEC). More recently, we had demonstrated that aldosterone modifies hemostasis via an upregulation of the protein-C receptor in HBMEC [18]. The results presented here demonstrate pleiotropic gene activation by the MCR-mediated action of aldosterone.
MATERIALS AND METHODS
HBMEC cultures, kindly provided by Pr. Pienta, were grown in M131 medium (Cascade Biologics, Portland, OR) supplemented with 10% Fetal Calf Serum (FCS) 100 IU/ml penicillin, 100 µg/ml streptomycin and 5% Microvascular growth supplement consisting of human basic fibroblast growth factor, heparin, human epidermal growth factor, and dibutyryl cyclic AMP (MVGS) obtained from Cascade Biologics, Inc.
In Vitro Angiogenesis Assay
The technique of Kern et al., (1983) [19] was adapted as follows: 1.5 x 105 HBMEC were cultured in 24 multiwell plates filled with M131 containing 5% FCS, 100 IU/ml penicillin, 100 µg/ml streptomycin and 5% MVGS. After 18 hours at 37°C, the cells were carefully washed 2 times with RPMI and detached with a non-enzymatic solution containing accutase (PAA, France). The effect of aldosterone, aldactone and eplerenone on capillary formation was analyzed on low matrigel base in a medium devoid of growth factor. To this end, 96-well plates were pre-coated with matrigel (Becton-Dickinson, Le Pont de Claix, France) that was allowed to polymerize for 1-2 hours at 37° C. Thereafter, 100 μl HBMEC (3.5 x 104 cells/ well) in M131, without FCS and MVGS, were seeded onto the matrigel-coated wells in presence of various concentrations and combinations of steroids. Following different incubation periods at 37° C, capillary tube formation was assessed using an inverted microscope fitted with a digital camera (Nikon-Diafot). The capillary network was quantitated by recording the number and length of connections between three or more capillary-like structures/field with the aid of the Archimed program, developed by Microvision [20].
Cell Migration
6 x 104 HBMEC were cultured in 24 well plates coated with 03% of gelatin in the presence of M131 containing 5% FCS and 5 % MVGS. After 18 h, the semi-confluent cells were dislodged by a cell scraper on a standardized surface. The effect of aldosterone (100 nM), eplerenone (400 nM), aldactone (400 nM), alone or in various combinations, on cell migration was quantitated by measuring the number of cells migrating to the wound edge after 6, 18, 27 and 48 h incubation in a medium containing only 2% FCS to avoid cell proliferation.
Cell Proliferation
6 x 104 HBMEC were cultured in 24 well plates in the presence of M 131 containing 10% FCS and aldosterone, eplerenone or aldactone in desired combinations. After 0, 18 and 36h, the cells were detached by trypsin and counted in the presence of Trypan blue.
Gene Array Analysis
The HBMEC cultures were exposed for 8 h to 100 nM aldosterone in M131, twenty five million cells were lysed in Trizol reagent and shipped to Memorec on dry ice. RNA was extracted and hybridized on PIQORtm microarrays, followed by linear amplification of RNA on oligo(dT)-T7 primed cDNA. Amplified RNA (aRNA) samples were quantitated by spectrophotometry and the sample quality was checked by gel electrophoresis (Bioanalyzer 2001, Agilent) as reported before [21]. The 1800 products in this microassay consisted of genes related to cytokines, monokines, growth factors, angiogenic effectors, cell metabolism, growth and malignant transformation. The experiment was repeated twice with four spots each time and is based on qPCR analysis.
Statistics
The student’s t test was used to assess the significance between various groups; the p values are shown in figure legends.
RESULTS
Inhibition of HBMEC Network by Aldosterone
Data in Fig. (1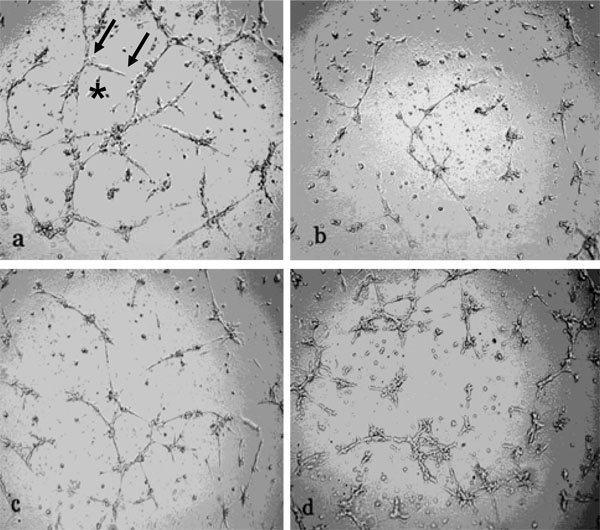 ) show decreased number of HBMEC networks and capillary lengths as a result of exposure to either 100 nM aldosterone (b), 400 nM eplerenone (c) or a combination of the agonist plus the antagonist (d), 18 h after the cells were seeded on low Matrigel support; in contrast, aldosterone had no effect on high matrigel support [16].
) show decreased number of HBMEC networks and capillary lengths as a result of exposure to either 100 nM aldosterone (b), 400 nM eplerenone (c) or a combination of the agonist plus the antagonist (d), 18 h after the cells were seeded on low Matrigel support; in contrast, aldosterone had no effect on high matrigel support [16].
 |
Fig. (1) Inhibition of HBMEC capillary length and abundance by aldosterone and eplerenone. HBMEC (30,000 cells) were exposed to 100
nM aldosterone (b), 400 nM eplerenone (c), or the agonist plus the antagonist (d), for 18 h and thereafter layered on Matrigel for 18 h.
Archimed program of Microvision was used to quantitate the capillary length (arrows in a) and the number of networks (*) compared to
control (a) in Fig. (1 |
Quantification by the Archimed program of Microvision revealed that the length of the capillaries decreased significantly from 17.000 mm in the control group (c) to 5.600 mm in the presence of 100 nM aldosterone, and to 11000 mm in the presence of eplerenone alone (Fig. 2A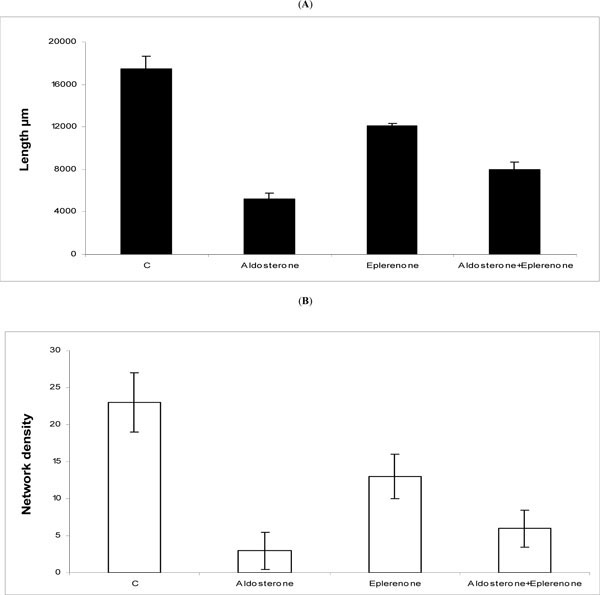 ). The potent inhibitory effect of aldosterone was partially attenuated by eplerenone such that the capillary length increased from, 5600 mm in the presence of aldosterone alone, to 8200 mm when the antagonist competed with the agonist. This suggests that both the agonist and the antagonist compete for MCR occupancy albeit the underlying mechanisms remain to be elucidated. Similarly, the number of capillary networks diminished significantly from 23 in the control group (c) to 3 and 12.5, in the presence of aldosterone and eplerenone, respectively (Fig. 2B). Again, eplerenone partially reversed the effect of aldosterone to raise the number of networks from 3 in the presence of the agonist alone to 6 when the antagonist was combined with the agonist (Fig. 2B). Similar results were obtained with aldactone when used in place of eplerenone (not shown).
). The potent inhibitory effect of aldosterone was partially attenuated by eplerenone such that the capillary length increased from, 5600 mm in the presence of aldosterone alone, to 8200 mm when the antagonist competed with the agonist. This suggests that both the agonist and the antagonist compete for MCR occupancy albeit the underlying mechanisms remain to be elucidated. Similarly, the number of capillary networks diminished significantly from 23 in the control group (c) to 3 and 12.5, in the presence of aldosterone and eplerenone, respectively (Fig. 2B). Again, eplerenone partially reversed the effect of aldosterone to raise the number of networks from 3 in the presence of the agonist alone to 6 when the antagonist was combined with the agonist (Fig. 2B). Similar results were obtained with aldactone when used in place of eplerenone (not shown).
 |
Fig. (2) Quantification of the number and the length of capillaries. The results, repeated three times, are expressed as the capillary length (Fig. 2A |
The Effect of Aldosterone and Eplerenone on HBMEC Migration
Data in Fig. (3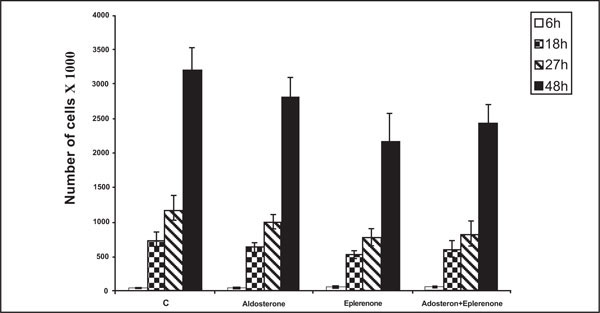 ) show time dependent HBMEC migration to the wound edge on the gelatine support. The number of migrating cells diminished from 3120 in the control group (c) to 2810, 2175 and 2420 in the presence of aldosterone, eplerenone, and aldosterone plus eplerenone, respectively, 48 h after a wound had been excised on the cell layer. In this test, aldosterone neither inhibited cell migration nor modified the significant inhibition observed with eplerenone. It would therefore appear that eplerenone possesses innate capacity to inhibit cell migration which is not dependent upon the antagonism of aldosterone action. Further experiments are needed to elucidate the action of eplerenone which assumes importance in view of the clinical use of the steroid [22, 23]. Interestingly, aldactone was even more potent than eplerenone in this context (data not shown) which may stem from the fact that it activates receptors for several steroid hormones (24 in contrast to the specificity of eplerenone for the MCR [22, 23].
) show time dependent HBMEC migration to the wound edge on the gelatine support. The number of migrating cells diminished from 3120 in the control group (c) to 2810, 2175 and 2420 in the presence of aldosterone, eplerenone, and aldosterone plus eplerenone, respectively, 48 h after a wound had been excised on the cell layer. In this test, aldosterone neither inhibited cell migration nor modified the significant inhibition observed with eplerenone. It would therefore appear that eplerenone possesses innate capacity to inhibit cell migration which is not dependent upon the antagonism of aldosterone action. Further experiments are needed to elucidate the action of eplerenone which assumes importance in view of the clinical use of the steroid [22, 23]. Interestingly, aldactone was even more potent than eplerenone in this context (data not shown) which may stem from the fact that it activates receptors for several steroid hormones (24 in contrast to the specificity of eplerenone for the MCR [22, 23].

Inhibition of HBMEC Proliferation by Aldosterone and Eplerenone
Data in Fig. (4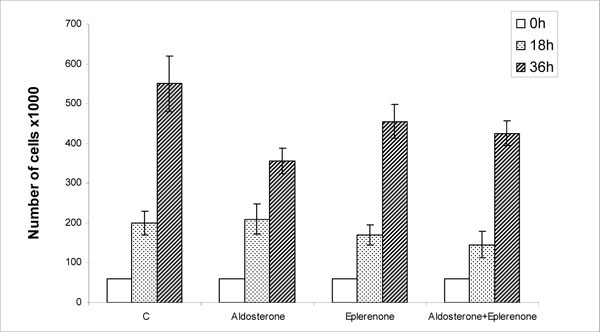 ) show that HBMEC proliferation, over the 36 h period, decreased from 550,000 in the control group to 380,000, 455,000 and 420,000 living cells in the presence of aldosterone, eplerenone and aldosterone plus eplerenone, respectively. Eplerenone was significantly less effective than aldosterone in inhibiting HBMEC proliferation and the agonist completely reversed the statistically insignificant inhibition observed with the antagonist alone. Aldactone was again more potent than eplerenone (not shown) possibly due to its affinity for various classes of steroid receptors [24].
) show that HBMEC proliferation, over the 36 h period, decreased from 550,000 in the control group to 380,000, 455,000 and 420,000 living cells in the presence of aldosterone, eplerenone and aldosterone plus eplerenone, respectively. Eplerenone was significantly less effective than aldosterone in inhibiting HBMEC proliferation and the agonist completely reversed the statistically insignificant inhibition observed with the antagonist alone. Aldactone was again more potent than eplerenone (not shown) possibly due to its affinity for various classes of steroid receptors [24].

Collectively, whereas eplerenone partially reversed both the angiogenic (Fig. 1) and growth (Fig. 4) inhibition by aldosterone the agonist did not influence cell migration at all (Fig. 3) which was effectively inhibited by eplerenone (Fig. 3). The reason(s) for these differences are not understood but could stem from the nature of gene(s) modified by MCR-mediated effect of aldosterone. In order to elucidate these relationships, a gene array analysis was performed to screen for those genes that could potentially respond to aldosterone. The 1800 products screened in this microassay represent genes that direct the synthesis of various cytokines, monokines, growth factors, angiogenic effectors, cell metabolism, cell growth and neoplasia.
Gene Modulation by Aldosterone
Data in Fig. (5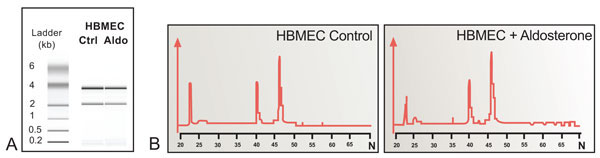 ) shows that the 28S/18S ratios were 1.72 and 1.85, respectively, for the control and aldosterone treated groups. As the samples did not show any additional degradation peaks in the low molecular range they were suitable for gene array screening. Data in Table 1 show upregulation of 20 mRNAs, along with the concurrent downregulation of 10 mRNAs in HBMEC exposed for 8 h to 100 nM aldosterone. In particular, the increase in mRNA for VEGF, CXCR-4, CDKN1A, Egr-1, EPCR, BCL-2, CD-24 and P-21 along with the decrease in the mRNA for NOD2, L1CAM, CD-83, ITGB-7, Cyclin-D, Cyclin-A, Decorin, GADD-135 and MMP-12, is in keeping with the accepted up and down regulation of these various factors in capillary formation. The table also provides the various synonyms in contemporary use to denote these various gene products. Altogether, 36 genes were up-regulated and 29 genes were down-regulated (details not shown).
) shows that the 28S/18S ratios were 1.72 and 1.85, respectively, for the control and aldosterone treated groups. As the samples did not show any additional degradation peaks in the low molecular range they were suitable for gene array screening. Data in Table 1 show upregulation of 20 mRNAs, along with the concurrent downregulation of 10 mRNAs in HBMEC exposed for 8 h to 100 nM aldosterone. In particular, the increase in mRNA for VEGF, CXCR-4, CDKN1A, Egr-1, EPCR, BCL-2, CD-24 and P-21 along with the decrease in the mRNA for NOD2, L1CAM, CD-83, ITGB-7, Cyclin-D, Cyclin-A, Decorin, GADD-135 and MMP-12, is in keeping with the accepted up and down regulation of these various factors in capillary formation. The table also provides the various synonyms in contemporary use to denote these various gene products. Altogether, 36 genes were up-regulated and 29 genes were down-regulated (details not shown).


DISCUSSION
In contrast to the classical view where aldosterone was dubbed a steroid specifically designed to regulate ion balance in the epithelial cells, it has now been highlighted as a key cardiovascular hormone where the steroid stimulates collagen deposition and left ventricular hypertrophy followed by ventricular remodeling [14, 25, 26]. Aldosterone is known to influence a number of important cardiac and vascular functions such as: inotropism, peripheral resistance, venous return, ventricular ejection, myocardial contractility, cyclooxygenase-2 and interleukin-6 [27] Na+-H exchanger isoform-1 [28], basolateral Na+/K+/ATPase [29, 30], reactive oxygen radicals [31], AP-1 [32], osteopontin [33], and the atrial natriuretic peptide gene product [34, 35].
The present study confirms and further extends our previous observations where the mineralocorticoid receptor-mediated action of steroid hormones induced the epithelial sodium channel in the endothelium [15]. The organization of endothelial cells into capillary-like networks was influenced by both aldosterone and the diuretic amiloride. The capillary diameter was also found dramatically increased, compared to the controls, as a result of aldosterone treatment [16]. As shown here, the underlying molecular events appear to consist of pleiotropic transactivation of genes that regulate capillary hemostasis in situ. Many other reports have variously demonstrated up and down regulation of c-kit ligand (stem cell factor, scf) [36], early growth response protein 1 (egr-1) [37], endothelial protein c receptor [18], cxcr4 [38], cdk-interacting protein 1-p21, [39], protein inhibitor of activated stat2 (pias2) [40], g1/s-specific cyclin d1 (prad1 oncogene [41, 42], growth arrest and dna damage inducible protein gadd153 [39], neural cell adhesion molecule l1 precursor (n-cam l1) (cd171 antigen) [43]. More recently, whereas one study has shown transcription regulation of many genes by aldosterone [44], in another report this steroid was shown to have no effect on transactivation [45] in human endothelium. Such discrepancies could stem from technical differences and cells used by various investigators.
The pleiotropic gene activation by aldosterone in the vascular endothelium, reported here, is in contrast with the classical view where the action of this steroid would be restricted to the upregulation of only a few genes that regulate the natriferic (Na+) action of the hormone in the epithelium [12-14]. Just as aldosterone exerts important cardiovascular function [14, 25, 26] so also this mineralocorticoid appears to assure the genesis, maintenance and renewal of the vascular architecture. Collectively, the vasculature indeed appears to function as an organ that is under the influence of many classes of cell signals and hormones.
DISCLOSURES
The authors have no financial conflicts of interest.
ACKNOWLEDGMENTS
We thank Pr. A. Therwath for many helpful discussions and “Retina France” for financial assistance.