- Home
- About Journals
-
Information for Authors/ReviewersEditorial Policies
Publication Fee
Publication Cycle - Process Flowchart
Online Manuscript Submission and Tracking System
Publishing Ethics and Rectitude
Authorship
Author Benefits
Reviewer Guidelines
Guest Editor Guidelines
Peer Review Workflow
Quick Track Option
Copyediting Services
Bentham Open Membership
Bentham Open Advisory Board
Archiving Policies
Fabricating and Stating False Information
Post Publication Discussions and Corrections
Editorial Management
Advertise With Us
Funding Agencies
Rate List
Kudos
General FAQs
Special Fee Waivers and Discounts
- Contact
- Help
- About Us
- Search




Current Chemical Genomics and Translational Medicine
(Discontinued)
ISSN: 2213-9885 ― Volume 12, 2018
A High Throughput Assay for Discovery of Small Molecules that Bind AMP-activated Protein Kinase (AMPK)
Sarah E Sinnett1, Jonathan Z Sexton2 , Jay E Brenman3, 4, *
Abstract
AMPK is a conserved heterotrimeric serine-threonine kinase that regulates anabolic and catabolic pathways in eukaryotes. Its central role in cellular and whole body metabolism makes AMPK a commonly proposed therapeutic target for illnesses characterized by abnormal energy regulation, including cancer and diabetes. Many AMPK modulators, however, produce AMPK-independent effects. To identify drugs that modulate AMPK activity independent of the canonical ATP-binding pocket found throughout the kinome, we designed a robust fluorescence-based high throughput screening assay biased toward the identification of molecules that bind the regulatory region of AMPK through displacement of MANT-ADP, a fluorescent ADP analog. Automated pin tools were used to rapidly transfer small molecules to a low volume assay mixture on 384-well plates. Prior to assay validation, we completed a full assay optimization to maximize the signal-to-background and reduce variability for robust detection of small molecules displacing MANT-ADP. After validation, we screened 13,120 molecules and identified 3 positive hits that dose-dependently inhibited the protein-bound signal of MANT-ADP in the presence of both full-length AMPK and the truncated “regulatory fragment” of AMPK, which is missing the kinase active site. The average Z’-factor for the screen was 0.55 and the compound confirmation rate was 60%. Thus, this fluorescence-based assay may be paired with in vitro kinase assays and cell-based assays to help identify molecules that selectively regulate AMPK with fewer off-target effects on other kinases.
Article Information
Identifiers and Pagination:
Year: 2013Volume: 7
First Page: 30
Last Page: 38
Publisher Id: CCGTM-7-30
DOI: 10.2174/2213988501307010030
Article History:
Received Date: 25/3/2013Revision Received Date: 25/4/2013
Acceptance Date: 25/4/2013
Electronic publication date: 3/9/2013
Collection year: 2013
open-access license: This is an open access article licensed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted, non-commercial use, distribution and reproduction in any medium, provided the work is properly cited.
* Address correspondence to this author at the Neuroscience Center and Department of Cell Biology and Physiology, UNC School of Medicine, Chapel Hill, NC 27599, USA, Tel: 919-843-3637, Fax: 919-966-9605, E-mail: brenman@med.unc.edu
| Open Peer Review Details | |||
|---|---|---|---|
| Manuscript submitted on 25-3-2013 |
Original Manuscript | A High Throughput Assay for Discovery of Small Molecules that Bind AMP-activated Protein Kinase (AMPK) | |
INTRODUCTION
AMP-activated protein kinase (AMPK) is a heterotrimeric serine-threonine kinase that regulates anabolic and catabolic pathways in eukaryotes [1Carling D, Thornton C, Woods A, Sanders MJ. AMP-activated protein kinase: new regulation, new roles? Biochem J 2012; 445(1): 11-27., 2Oakhill JS, Scott JW, Kemp BE. AMPK functions as an adenylate charge-regulated protein kinase Trends Endocrinol Metab 2012; 23(3): 125-32.]. AMPK’s ubiquitous expression and central role in cellular metabolism have made it a commonly proposed therapeutic target for disorders characterized by abnormal energy regulation, including cancer and diabetes [1Carling D, Thornton C, Woods A, Sanders MJ. AMP-activated protein kinase: new regulation, new roles? Biochem J 2012; 445(1): 11-27.]. Drugs that specifically target AMPK could have immense translational value. The earliest AMPK modulators, AICAR and Compound C, were not specifically developed for AMPK and have off-target effects [3Kim M, Tian R. Targeting AMPK for cardiac protection: opportunities and challenges J Mol Cell Cardiol 2011; 51(4): 548-3., 4Zhou G, Myers R, Li Y, et al. Role of AMP-activated protein kinase in mechanism of metformin action J Clin Invest 2001; 108(8): 1167-74.]. Compound C, which was identified in a library screen for AMPK inhibitors, binds the canonical ATP-binding catalytic site in numerous kinases [3Kim M, Tian R. Targeting AMPK for cardiac protection: opportunities and challenges J Mol Cell Cardiol 2011; 51(4): 548-3.-5Handa N, Takagi T, Saijo S, et al. Structural basis for compound C inhibition of the human AMP-activated protein kinase alpha2 subunit kinase domain Acta Crystallogr D Biol Crystallogr 2011; 67(Pt 5): 480-7.]. Strategies designed to identify AMPK modulators that target AMPK-unique protein domains, therefore, have the theoretical potential to identify molecules that are more selective than those that bind the canonical ATP-binding catalytic site found throughout the kinome.
The mammalian AMPK trimer is composed of one catalytic subunit (α1 or α2) and two regulatory subunits (β1 or β2 and γ1, γ2, or γ3) [1Carling D, Thornton C, Woods A, Sanders MJ. AMP-activated protein kinase: new regulation, new roles? Biochem J 2012; 445(1): 11-27., 2Oakhill JS, Scott JW, Kemp BE. AMPK functions as an adenylate charge-regulated protein kinase Trends Endocrinol Metab 2012; 23(3): 125-32.]. AMPK-γ has four potential nucleotide-binding pockets, of which one (Site 2) is constitutively unoccupied [6Xiao B, Heath R, Saiu P, et al. Structural basis for AMP binding to mammalian AMP-activated protein kinase Nature 2007; 449(7161): 496-500.]. Previous studies have identified several modes of AMPK regulation: (1) Phosphorylation of threonine 172 (T172) on AMPK-α activates AMPK; (2) Binding of AMP to AMPK-γ (Site 1) allosterically increases activity of p-AMPK by 2 to 5-fold; and (3) Binding of AMP or ADP to AMPK-γ (Site 3) inhibits dephosphorylation of p-T172 by phosphatases [2Oakhill JS, Scott JW, Kemp BE. AMPK functions as an adenylate charge-regulated protein kinase Trends Endocrinol Metab 2012; 23(3): 125-32., 7Xiao B, Sanders MJ, Underwood E, et al. Structure of mammalian AMPK and its regulation by ADP Nature 2011; 472(7342): 230-3.]. AMP and ADP have also been shown to promote the phosphorylation of AMPK, but only when AMPK-β is myristoylated [1Carling D, Thornton C, Woods A, Sanders MJ. AMP-activated protein kinase: new regulation, new roles? Biochem J 2012; 445(1): 11-27., 8Oakhill JS, Chen ZP, Scott JW, et al. beta-Subunit myristoylation is the gatekeeper for initiating metabolic stress sensing by AMP-activated protein kinase (AMPK) Proc Natl Acad Sci USA 2010; 107(45): 19237-41.-10Oakhill JS, Steel R, Chen ZP, et al. AMPK is a direct adenylate charge-regulated protein kinase Science 2011; 332(6036): 1433-5.]. Site 4 has been proposed to bind AMP in a non-exchangeable manner [6Xiao B, Heath R, Saiu P, et al. Structural basis for AMP binding to mammalian AMP-activated protein kinase Nature 2007; 449(7161): 496-500., 7Xiao B, Sanders MJ, Underwood E, et al. Structure of mammalian AMPK and its regulation by ADP Nature 2011; 472(7342): 230-3.]. Recent co-crystallization experiments, however, have shown that nucleotide exchange with ATP is possible, though perhaps not physiological [9Chen L, Wang J, Zhang YY, et al. AMP-activated protein kinase undergoes nucleotide-dependent conformational changes Nat Struct Mol Biol 2012; 19(7): 716-8.]. Although some researchers have disagreed with the designation of Site 3 as the dephosphorylation inhibition site, most AMPK researchers agree that AMPK-γ has two primary regulatory nucleotide-binding sites that, together, allosterically increase and sustain activity of p-AMPK [7Xiao B, Sanders MJ, Underwood E, et al. Structure of mammalian AMPK and its regulation by ADP Nature 2011; 472(7342): 230-3., 9Chen L, Wang J, Zhang YY, et al. AMP-activated protein kinase undergoes nucleotide-dependent conformational changes Nat Struct Mol Biol 2012; 19(7): 716-8., 10Oakhill JS, Steel R, Chen ZP, et al. AMPK is a direct adenylate charge-regulated protein kinase Science 2011; 332(6036): 1433-5.].
Historically, AMP has been thought to be the primary allosteric regulator of p-AMPK [1Carling D, Thornton C, Woods A, Sanders MJ. AMP-activated protein kinase: new regulation, new roles? Biochem J 2012; 445(1): 11-27., 11Sim AT, Hardie DG. The low activity of acetyl-CoA carboxylase in basal and glucagon-stimulated hepatocytes is due to phosphorylation by the AMP-activated protein kinase and not cyclic AMP-dependent protein kinase FEBS Lett 1988; 233(2): 294-8.]. However, ADP’s greater cytosolic abundance and ability to inhibit dephosphorylation of p-T172 have led an increasing number of investigators to postulate that ADP is the primary allosteric AMPK regulator in vivo [1Carling D, Thornton C, Woods A, Sanders MJ. AMP-activated protein kinase: new regulation, new roles? Biochem J 2012; 445(1): 11-27.]. The discovery of ADP’s regulatory role was foreshadowed nearly a decade ago, when Chen and colleagues demonstrated a correlation between ADP levels and AMPK activity measured from human skeletal muscle before and after intense exercise [12Chen ZP, Stephens TJ, Murthy S, et al. Effect of exercise intensity on skeletal muscle AMPK signaling in humans Diabetes 2003; 52(9): 2205-12.]. Furthermore, these investigators demonstrated that the rise in ADP dwarfed the slight increase in AMP in human exercised muscle by 19-fold [12Chen ZP, Stephens TJ, Murthy S, et al. Effect of exercise intensity on skeletal muscle AMPK signaling in humans Diabetes 2003; 52(9): 2205-12.]. Given the similar affinities of AMP and ADP for the exchangeable AMPK-γ sites (2.5 and 1.5 μM at Site 1, respectively; 80 and 50 μM at Site 3, respectively), it is more likely that ADP is the primary regulatory nucleotide of AMPK in vivo [7Xiao B, Sanders MJ, Underwood E, et al. Structure of mammalian AMPK and its regulation by ADP Nature 2011; 472(7342): 230-3.].
Although AMPK-α and AMPK-γ can both bind adenine nucleotides, the binding sites on these subunits can have differential affinities for ligands. For example, the non-specific kinase inhibitor staurosporine binds only the single AMPK-α canonical catalytic site [7Xiao B, Sanders MJ, Underwood E, et al. Structure of mammalian AMPK and its regulation by ADP Nature 2011; 472(7342): 230-3.]. In addition, the environment-sensitive fluorophore 2'/3'-O-(N-methylanthraniloyl)-ATP (MANT-ATP) binds two sites on both truncated and full-length AMPK, even though full-length AMPK contains three potentially exchangeable nucleotide-binding sites [6Xiao B, Heath R, Saiu P, et al. Structural basis for AMP binding to mammalian AMP-activated protein kinase Nature 2007; 449(7161): 496-500.]. The heterotrimeric truncated AMPK (in contrast to full-length AMPK trimer) lacks the canonical ATP-binding site found throughout the kinome and is termed the AMPK “regulatory fragment” [6Xiao B, Heath R, Saiu P, et al. Structural basis for AMP binding to mammalian AMP-activated protein kinase Nature 2007; 449(7161): 496-500., 7Xiao B, Sanders MJ, Underwood E, et al. Structure of mammalian AMPK and its regulation by ADP Nature 2011; 472(7342): 230-3., 13Oakhill JS, Scott JW, Kemp BE. Structure and function of AMP-activated protein kinase Acta Physiol (Oxf) 2009; 196(1): 3-14.]. The differential affinities of binding sites on AMPK-α and AMPK-γ for various ligands suggests that it may be possible to identify novel AMPK modulators that bind the regulatory fragment but not the canonical ATP-binding site found throughout most of the kinome.
We designed a rapid fluorescence-based high throughput assay to identify small molecules that would displace MANT-ADP bound to full-length AMPK. MANT-ADP’s fluorescence increases after binding to AMPK, and competitive displacement of bound MANT-ADP by the primary regulatory nucleotide ADP reverses this increase in fluorescence. AMPK has previously been shown to have two MANT-adenine nucleotide-binding sites on the regulatory fragment [6Xiao B, Heath R, Saiu P, et al. Structural basis for AMP binding to mammalian AMP-activated protein kinase Nature 2007; 449(7161): 496-500., 14Saiu P. Structural and functional studies on nucleotide binding to AMP-activated protein kinase. London: University College London 2010. [Accessed March 2013]; Available: http://discovery.ucl.ac.uk/64-5676/1/645676.pdf]. This assay could therefore identify molecules that compete with MANT-ADP for binding to Site 1 or Site 3 – the exchangeable sites on AMPK-γ. Theoretically, the assay could also identify molecules that bind to an unknown allosteric site, provided the binding of the small molecule and MANT-ADP are mutually exclusive. After screening 13,120 small molecules at less than 5 µM, we identified 3 molecules that dose-dependently inhibit MANT-ADP fluorescence in the presence of full-length AMPK (the “primary assay/screen”). We also developed a similar secondary assay using the trimeric AMPK regulatory fragment lacking the kinase domain (the “secondary assay/screen”). All 3 positive hits dose-dependently inhibited MANT-ADP fluorescence in the regulatory fragment assay as well.
MATERIALS AND METHODOLOGY
Materials
STK740822 (Vitas-M Laboratory, Ltd.), STL035166 (Vitas-M Laboratory, Ltd.), and Z64358107 (Enamine, Ltd.) were re-purchased in powder form and then dissolved in DMSO for dose response studies. ADP, lysozyme, and 1% EDTA-free protease inhibitor cocktail were purchased from Sigma-Aldrich. MANT-ADP was purchased from Life Technologies. Cobalt-based immobilized metal affinity chromatography (IMAC) resin was purchased from Clontech Laboratories, Inc.
Protein Purification
Tricistronic vectors encoding full-length AMPK (rat His-α1, β1, γ1) and the AMPK regulatory fragment (rat His-α1, 396-548; human β2, 187-272; rat γ1) were gifts from Dr. Uwe Schlattner and Dr. Steve Gamblin, respectively. Vectors were transformed into Rosetta cells and individual colonies were then incubated at 37 °C in overnight starter cultures containing antibiotics. Overnight cultures were amplified in auto-inducible media and shaken for two days at room temperature. Induced cultures were pelleted, washed twice in 0.9% NaCl, and then sonicated (40% intensity, 30 seconds, 3 times with 2 minute intervals) at 4 °C in lysis buffer containing 50 mM Tris-HCl (pH 8), 100 mM NaCl, 0.75 mg/mL lysozyme, 0.1% Triton X100, and 1% EDTA-free protease inhibitor cocktail. Lysates were centrifuged and supernatants were batch-bound onto cobalt-based IMAC resin. Resin was washed three times in buffer containing 0.01% Triton X100 (first wash only), 50 mM Tris-HCl (pH 8), 100 mM NaCl, and 2 mM imidazole. Washed resin was loaded onto a column prior to elution in 50 mM Tris-HCl (pH 8), 100 mM NaCl, and 500 mM imidazole. Eluates were concentrated on either 30 kDa (for the regulatory fragment) or 50 kDa (for full-length AMPK) size exclusion centrifugal filters at 4 °C. Concentrates were resuspended and concentrated twice more in 50 mM Tris-HCl (pH 8) prior to storage at -80 °C.
Assay Assembly
Except where otherwise noted, assays were performed with the following conditions: 0.5 μM full-length AMPK (130 kDa) or 0.5 μM regulatory fragment (66.7 kDa), 0.1 μM MANT-ADP, 10 mM Tris-HCl (pH 8), 0.45% DMSO (vehicle and negative control), and 5 μM ADP (positive control). Automated assays were assembled in two steps. First, a NanoQuot (BioTek) was used to dispense 11 μL of master mix (protein, MANT-ADP, and Tris-HCl) per well in black, low volume 384-well plates (Corning-3676). Next, a Biomek NX workstation (Beckman-Coulter, Brea, CA) equipped with pin tools (VP Scientific, San Diego CA) was used to transfer 50 nL of controls (columns 1, 2, 23, and 24) and small molecules (4.5 μM final). Controls and library molecules were transferred from Axygen 384-well rigid PCR plates (Cat no. 321-67-051) at 221X concentration for single-concentration screening. Plates were shaken for 10 minutes prior to reading.
Fluorescence Measurements
Fluorescence emission spectra of MANT-ADP were collected using a PerkinElmer EnSpire plate reader with an excitation wavelength of 360 nm. Background fluorescence from protein was subtracted from the raw data prior to plotting MANT-ADP’s fluorescence in the presence of protein. Fluorescence detection for assay development and screening was performed on either a BMG Pherastar Plus (360/10 excitation and 460/10 emission filters) or a PerkinElmer EnVision (355/40 excitation and 460/25 emission filters), depending on equipment availability. Relative fluorescence units (RFUs) were recorded at room temperature.
Calculation of Z’
Z’ was calculated using the formula: Z’ = 1- ((3(σvehicle + σADP))/(μvehicle – μADP)) [15Zhang JH, Chung TD, Oldenburg KR. A simple statistical parameter for use in evaluation and validation of high throughput screening assays J Biomol Screen 1999; 4(2): 67-73.].
Small Molecule Library
The Center for Integrative Chemical Biology and Drug Discovery (CICBDD) assembled a commercially-available small molecule library from multiple sources comprised of molecules that have structural similarities to known kinase inhibitors and/or have been shown to bind kinases in silico [16Hutti JE, Porter MA, Cheely AW, et al. Development of a high-throughput assay for identifying inhibitors of TBK1 and IKKepsilon PLoS ONE 2012; 7(7): e41494., 17Peterson EJ, Janzen WP, Kireev D, Singleton SF. High-throughput screening for RecA inhibitors using a transcreener adenosine 5'-O-diphosphate assay Assay Drug Dev Technol 2012; 10(3): 260-8.]. Library molecules were stored at 1 mM concentration in DMSO. All of the molecules comply with Lipinski’s rules [18Lipinski CA, Lombardo F, Dominy BW, Feeney PJ. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings Adv Drug Deliv Rev 1997; 23(1-3): 3-25.].
Screening Data Analysis
ScreenAble (ScreenAble Solutions, Chapel Hill, NC) high throughput screening software was utilized for merging screening data and chemical structure, for statistical design of experiments, and hit selection. GraphPad Prism was used for non-linear regression analysis of dose response data.
RESULTS
The overall goal of this study was to identify novel small molecules that preferentially bind the regulatory region of AMPK. To achieve this goal, we designed a fluorescence-based assay using purified His-tagged AMPK trimer and a fluorescent analog of AMPK’s primary regulatory nucleotide, ADP (Fig. S1). Optimizing lysis conditions and switching from IPTG-inducible media to auto-inducible media increased the yield of purified protein from 3 mgs to 20 mgs per liter of culture, greater than previously published yields for AMPK heterotrimers [19Neumann D, Woods A, Carling D, Wallimann T, Schlattner U. Mammalian AMP-activated protein kinase: functional, heterotrimeric complexes by co-expression of subunits in Escherichia coli Protein Expr Purif 2003; 30(2): 230-7.-21Riek U, Scholz R, Konarev P, et al. Structural properties of AMP-activated protein kinase: dimerization, molecular shape, and changes upon ligand binding J Biol Chem 2008; 283(26): 18331-43.]. This improved AMPK expression method supplied ample protein for designing and optimizing assays, screening libraries, and confirming hits in secondary assays.
After a thorough review of the available fluorescent nucleotide analogs and assay technologies, fluorescent ADP analogs were chosen because of the known physiologic role of ADP interaction with AMPK [7Xiao B, Sanders MJ, Underwood E, et al. Structure of mammalian AMPK and its regulation by ADP Nature 2011; 472(7342): 230-3., 10Oakhill JS, Steel R, Chen ZP, et al. AMPK is a direct adenylate charge-regulated protein kinase Science 2011; 332(6036): 1433-5.]. Both MANT-labeled and trinitrophenylated (TNP) nucleotide analogs are environment-sensitive probes whose fluorescence increases upon binding to nucleotide-binding sites on protein (Fig. 1A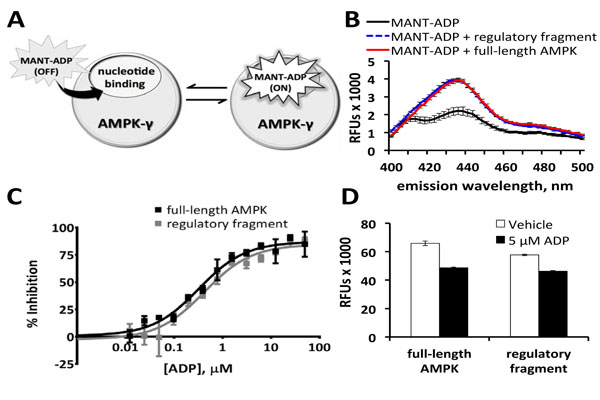 ) [6Xiao B, Heath R, Saiu P, et al. Structural basis for AMP binding to mammalian AMP-activated protein kinase Nature 2007; 449(7161): 496-500., 14Saiu P. Structural and functional studies on nucleotide binding to
AMP-activated protein kinase. London: University College
London 2010. [Accessed March 2013]; Available:
http://discovery.ucl.ac.uk/64-5676/1/645676.pdf, 22Guarnieri MT, Blagg BS, Zhao R. A high-throughput TNP-ATP displacement assay for screening inhibitors of ATP-binding in bacterial histidine kinases Assay Drug Dev Technol 2011; 9(2): 174-83.]. Fluorescence emission spectra were recorded for MANT-ADP and TNP-ADP (0.1 μM each) in the presence and absence of protein (Figs. 1B,
S2). Only MANT-ADP fluorescence increased upon addition of full-length AMPK and the regulatory fragment (Fig. 1B). MANT-ADP was therefore selected for detection of small molecules binding to the regulatory fragment of AMPK. The general assay principle is to pre-load AMPK with MANT-ADP, causing an increase in MANT-ADP fluorescence. If a small molecule can displace MANT-ADP from the binding site, MANT-ADP will be ejected and will have a substantial decrease in fluorescence, thereby indicating a positive result for the test compound. Using excess AMPK (0.5 µM protein with 0.1 µM MANT-ADP) helped maximize the number of MANT-ADP molecules bound to protein and thus helped maximize the protein-bound fluorescent signal in the absence of the competitive positive control ADP.
) [6Xiao B, Heath R, Saiu P, et al. Structural basis for AMP binding to mammalian AMP-activated protein kinase Nature 2007; 449(7161): 496-500., 14Saiu P. Structural and functional studies on nucleotide binding to
AMP-activated protein kinase. London: University College
London 2010. [Accessed March 2013]; Available:
http://discovery.ucl.ac.uk/64-5676/1/645676.pdf, 22Guarnieri MT, Blagg BS, Zhao R. A high-throughput TNP-ATP displacement assay for screening inhibitors of ATP-binding in bacterial histidine kinases Assay Drug Dev Technol 2011; 9(2): 174-83.]. Fluorescence emission spectra were recorded for MANT-ADP and TNP-ADP (0.1 μM each) in the presence and absence of protein (Figs. 1B,
S2). Only MANT-ADP fluorescence increased upon addition of full-length AMPK and the regulatory fragment (Fig. 1B). MANT-ADP was therefore selected for detection of small molecules binding to the regulatory fragment of AMPK. The general assay principle is to pre-load AMPK with MANT-ADP, causing an increase in MANT-ADP fluorescence. If a small molecule can displace MANT-ADP from the binding site, MANT-ADP will be ejected and will have a substantial decrease in fluorescence, thereby indicating a positive result for the test compound. Using excess AMPK (0.5 µM protein with 0.1 µM MANT-ADP) helped maximize the number of MANT-ADP molecules bound to protein and thus helped maximize the protein-bound fluorescent signal in the absence of the competitive positive control ADP.

ADP, which competitively binds to Site 1 and Site 3 on AMPK-γ, inhibited the increase in MANT-ADP fluorescence with IC50s of 0.4 μM and 0.3 μM for the regulatory fragment and full-length AMPK, respectively (Fig. 1C). For the ADP dose responses, replicates containing MANT-ADP with no protein were used as positive controls for 100% inhibition of MANT-ADP’s protein-bound fluorescent signal. Although the signal-to-background ratio was less than 2-fold (Figs. 1B, 1D), the assay’s Z’-factor was greater than 0.6 (Fig. 1D), indicating that the assay was robust enough for high throughput screening. At an emission wavelength of 460 nm, full-length AMPK consistently provided a slightly larger assay window, usually resulting in higher Z’-factors (Fig. 1D). The small molecule library, therefore, was screened against full-length AMPK. Positive hits were confirmed against the regulatory fragment in subsequent secondary screens. Aside from a small difference in assay window, truncation of AMPK-α1 and AMPK-β2 did not appear to significantly disrupt interactions among AMPK-γ1, MANT-ADP, and ADP.
Prior to screening, assay conditions were optimized by testing high and low concentrations of several reagents in a design of experiments study using ScreenAble software (ScreenAble Solutions, Chapel Hill, NC). Previous studies have shown that affinities of adenine nucleotides for AMPK decrease with increasing ionic strength [6Xiao B, Heath R, Saiu P, et al. Structural basis for AMP binding to mammalian AMP-activated protein kinase Nature 2007; 449(7161): 496-500., 14Saiu P. Structural and functional studies on nucleotide binding to
AMP-activated protein kinase. London: University College
London 2010. [Accessed March 2013]; Available:
http://discovery.ucl.ac.uk/64-5676/1/645676.pdf]. In agreement with published data, the greatest MANT-ADP fluorescence was observed with a low concentration of Tris-HCl (pH 8) and 0 µM NaCl (Fig. 2A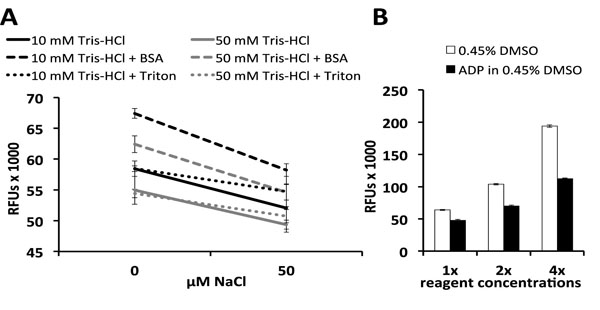 ). Triton, which is often used to prevent adsorption of target proteins onto plastic, had no effect on MANT-ADP fluorescence in the presence of 0 µM NaCl (Fig. 2A) [23Simpson RJ. Stabilization of proteins for storage Cold Spring Harb Protoc pdb top 2010; 2010(5) pdb top79]. The sacrificial protein BSA did increase fluorescence (Fig. 2A), but this was due in part to interactions between BSA and MANT-ADP. In the absence of AMPK, MANT-ADP’s fluorescence still increased upon addition of BSA, even after subtracting BSA’s autofluorescence from the raw data (Fig.
S3A). It is possible that MANT-ADP binds non-specifically to BSA, thus decreasing the pool of MANT-ADP molecules that can bind to AMPK and consequently decreasing the assay window between vehicle and ADP-treated control groups (Fig.
S3B). Because BSA decreased the assay window and Z’-factor, we decided to exclude BSA from our optimized assay conditions (Fig.
S3B). Optimized buffer conditions yielded a Z’-factor ≥ 0.6 with an assay window that increased linearly with protein and MANT-ADP concentrations (Fig. 2B). Instead of increasing AMPK and MANT-ADP concentrations to maximize the assay window, we decided to optimize the assay with low reagent concentrations (0.5 μM AMPK and 0.1 μM MANT-ADP) to ensure sensitivity for small molecule binding, as micromolar concentrations of AMPK would severely limit the theoretical maximum inhibition due to the stoichiometry of enzyme to small molecule.
). Triton, which is often used to prevent adsorption of target proteins onto plastic, had no effect on MANT-ADP fluorescence in the presence of 0 µM NaCl (Fig. 2A) [23Simpson RJ. Stabilization of proteins for storage Cold Spring Harb Protoc pdb top 2010; 2010(5) pdb top79]. The sacrificial protein BSA did increase fluorescence (Fig. 2A), but this was due in part to interactions between BSA and MANT-ADP. In the absence of AMPK, MANT-ADP’s fluorescence still increased upon addition of BSA, even after subtracting BSA’s autofluorescence from the raw data (Fig.
S3A). It is possible that MANT-ADP binds non-specifically to BSA, thus decreasing the pool of MANT-ADP molecules that can bind to AMPK and consequently decreasing the assay window between vehicle and ADP-treated control groups (Fig.
S3B). Because BSA decreased the assay window and Z’-factor, we decided to exclude BSA from our optimized assay conditions (Fig.
S3B). Optimized buffer conditions yielded a Z’-factor ≥ 0.6 with an assay window that increased linearly with protein and MANT-ADP concentrations (Fig. 2B). Instead of increasing AMPK and MANT-ADP concentrations to maximize the assay window, we decided to optimize the assay with low reagent concentrations (0.5 μM AMPK and 0.1 μM MANT-ADP) to ensure sensitivity for small molecule binding, as micromolar concentrations of AMPK would severely limit the theoretical maximum inhibition due to the stoichiometry of enzyme to small molecule.

Since many small molecule libraries use DMSO as a solvent, the DMSO tolerance of the optimized assay was determined prior to screening. The assay yielded a robust Z’ across a range of DMSO concentrations, with no statistical difference observed among controls treated with 0-2% DMSO (Fig. S4). This DMSO tolerance is especially important for automated assay assembly, in which the final DMSO concentration may vary depending on the type of robotic dispenser used for transferring small molecules. The DMSO tolerance also allows flexibility in starting concentrations of library plate stocks.
For library screening, a NanoQuot and a Biomek NX workstation were used to dispense the assay master mix and small molecules, respectively. To validate the automated dispensers, the plate-to-plate coefficient of variation (CV) was calculated for the control groups on 4 plates (< 4% CV between plates; average Z’-factor = 0.6) (Fig. 3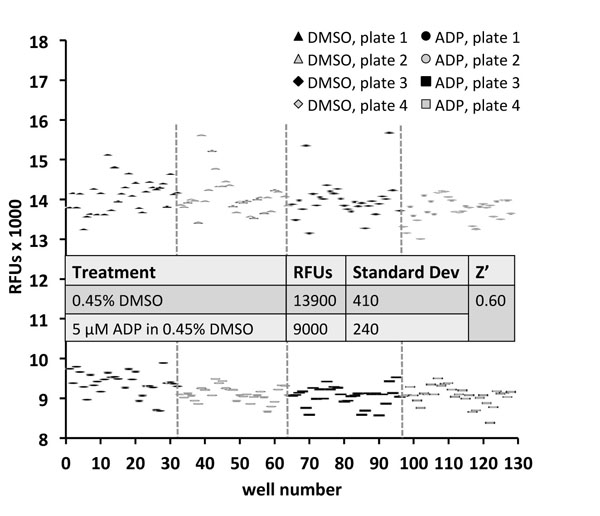 ). Fluorescence detection was performed on a BMG Pherastar Plus, which produced lower signals than the PerkinElmer EnVision used for other figures (Figs. 1D, 2). The fold-difference between MANT-ADP’s protein-bound and unbound signals, however, was similar for both detectors. After validating minimal plate-to-plate variation, 13,120 molecules from a small molecule library were screened over the course of several days. Five positive hits (0.04% primary hit rate) were identified, each of which inhibited MANT-ADP fluorescence by more than 50% (Fig. 4A
). Fluorescence detection was performed on a BMG Pherastar Plus, which produced lower signals than the PerkinElmer EnVision used for other figures (Figs. 1D, 2). The fold-difference between MANT-ADP’s protein-bound and unbound signals, however, was similar for both detectors. After validating minimal plate-to-plate variation, 13,120 molecules from a small molecule library were screened over the course of several days. Five positive hits (0.04% primary hit rate) were identified, each of which inhibited MANT-ADP fluorescence by more than 50% (Fig. 4A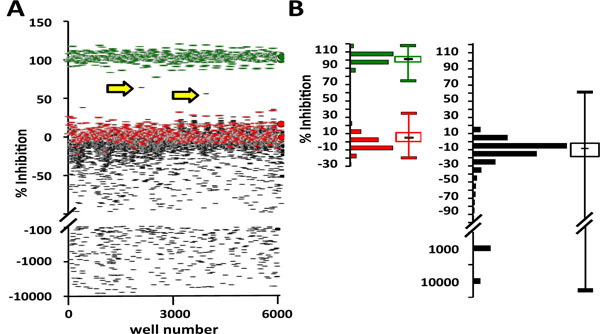 ). Autofluorescent molecules increased the total fluorescent signal and are shown on the scatter plot as having large negative binding activity (Fig. 4A). Data for DMSO controls and library molecules had similar distributions, with the average compound binding activity close to 0% inhibition of MANT-ADP fluorescence (Fig. 4B). The large number of autofluorescent molecules, which decreased the efficiency of screening, may explain the low primary hit rate. It is possible that some of the autofluorescent molecules are false negatives that inhibit binding of MANT-ADP but are masked by their autofluorescence. After initial screening, positive hits were re-purchased in powder form, dissolved in DMSO and re-tested for inhibition of MANT-ADP fluorescence. Three of the 5 positive hits (60% confirmation rate) dose-dependently inhibited MANT-ADP fluorescence in the presence of full-length AMPK (Fig. 5
). Autofluorescent molecules increased the total fluorescent signal and are shown on the scatter plot as having large negative binding activity (Fig. 4A). Data for DMSO controls and library molecules had similar distributions, with the average compound binding activity close to 0% inhibition of MANT-ADP fluorescence (Fig. 4B). The large number of autofluorescent molecules, which decreased the efficiency of screening, may explain the low primary hit rate. It is possible that some of the autofluorescent molecules are false negatives that inhibit binding of MANT-ADP but are masked by their autofluorescence. After initial screening, positive hits were re-purchased in powder form, dissolved in DMSO and re-tested for inhibition of MANT-ADP fluorescence. Three of the 5 positive hits (60% confirmation rate) dose-dependently inhibited MANT-ADP fluorescence in the presence of full-length AMPK (Fig. 5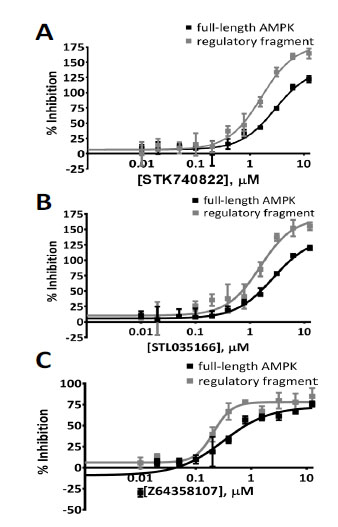 , Table 1). These 3 hits also dose-dependently inhibited fluorescence in the presence of the regulatory fragment (Fig. 5, Table 1). Like the primary assay using full-length AMPK, the secondary assay was performed in buffer that had low ionic strength and was assembled in the same manner, with controls and positive hits added to a master mix of regulatory fragment and MANT-ADP.
, Table 1). These 3 hits also dose-dependently inhibited fluorescence in the presence of the regulatory fragment (Fig. 5, Table 1). Like the primary assay using full-length AMPK, the secondary assay was performed in buffer that had low ionic strength and was assembled in the same manner, with controls and positive hits added to a master mix of regulatory fragment and MANT-ADP.



DISCUSSION
AMPK’s central role in metabolism makes it an appealing therapeutic target for numerous illnesses characterized by abnormalities in energy regulation, such as diabetes or cancer [1Carling D, Thornton C, Woods A, Sanders MJ. AMP-activated protein kinase: new regulation, new roles? Biochem J 2012; 445(1): 11-27.]. The current widely used AMPK modulators, however, were not specifically developed for AMPK and have off-target effects [3Kim M, Tian R. Targeting AMPK for cardiac protection: opportunities and challenges J Mol Cell Cardiol 2011; 51(4): 548-3., 4Zhou G, Myers R, Li Y, et al. Role of AMP-activated protein kinase in mechanism of metformin action J Clin Invest 2001; 108(8): 1167-74.]. Compound C, which was identified in a library screen for AMPK inhibitors, binds the catalytic ATP-binding site and has been shown to be a more potent inhibitor of numerous other kinases [3Kim M, Tian R. Targeting AMPK for cardiac protection: opportunities and challenges J Mol Cell Cardiol 2011; 51(4): 548-3.-5Handa N, Takagi T, Saijo S, et al. Structural basis for compound C inhibition of the human AMP-activated protein kinase alpha2 subunit kinase domain Acta Crystallogr D Biol Crystallogr 2011; 67(Pt 5): 480-7., 24Machrouhi F, Ouhamou N, Laderoute K, et al. The rational design of a novel potent analogue of the 5'-AMP-activated protein kinase inhibitor compound C with improved selectivity and cellular activity Bioorg Med Chem Lett 2010; 20(22): 6394-9.]. Furthermore, Compound C’s selectivity may be over-estimated in AMPK literature, as many papers fail to mention Compound C’s alternate identity as the bone morphogenetic protein receptor modulator, dorsomorphin [25Yu PB, Hong CC, Sachidanandan C, et al. Dorsomorphin inhibits BMP signals required for embryogenesis and iron metabolism Nat Chem Biol 2008; 4(1): 33-41.]. In contrast to molecules that bind the canonical catalytic ATP-binding pocket found throughout the kinome, molecules that bind the regulatory region of AMPK may be more selective and could be extremely useful as either drugs or as tool compounds for analysis of AMPK function in vivo. It is important to note that both Site 1 and Site 3 bind AMP, ADP, and ATP [7Xiao B, Sanders MJ, Underwood E, et al. Structure of mammalian AMPK and its regulation by ADP Nature 2011; 472(7342): 230-3.]. Small molecules that bind the regulatory region of AMPK may, therefore, include ATP-competitive ligands. The ideal discovery would be that of an ATP-competitive modulator that binds Site 1 and/or Site 3, but not the kinase active site. The preferential binding of staurosporine to the kinase active site indicates differential affinities of these three ATP-binding sites for small molecules [7Xiao B, Sanders MJ, Underwood E, et al. Structure of mammalian AMPK and its regulation by ADP Nature 2011; 472(7342): 230-3.].
To identify molecules that bind the regulatory region of AMPK, we designed a fluorescence-based assay using readily purified His-tagged protein and commercially available MANT-ADP, an analog of AMPK’s primary regulatory nucleotide [1Carling D, Thornton C, Woods A, Sanders MJ. AMP-activated protein kinase: new regulation, new roles? Biochem J 2012; 445(1): 11-27.]. Binding of MANT-labeled analogs to nucleotide-binding sites on protein can be easily detected by an increase in fluorescence [6Xiao B, Heath R, Saiu P, et al. Structural basis for AMP binding to mammalian AMP-activated protein kinase Nature 2007; 449(7161): 496-500.]. No crystal structures of AMPK with MANT-ADP have been published, but structures of the regulatory fragment soaked with MANT-AMP indicate binding at Sites 1 and 3 [14Saiu P. Structural and functional studies on nucleotide binding to AMP-activated protein kinase. London: University College London 2010. [Accessed March 2013]; Available: http://discovery.ucl.ac.uk/64-5676/1/645676.pdf]. Because full-length AMPK provided a slightly larger assay window in preliminary experiments, we decided to use full-length AMPK for our primary screen and the regulatory fragment for our secondary screen. Although full-length AMPK has three available nucleotide- binding sites (one on AMPK-α and two on AMPK-γ), fluorescence studies have shown that MANT-ATP binds only two sites on both the regulatory fragment and full-length AMPK [6Xiao B, Heath R, Saiu P, et al. Structural basis for AMP binding to mammalian AMP-activated protein kinase Nature 2007; 449(7161): 496-500.]. Because our assay measures MANT-ADP fluorescence and not enzyme activity, the assay could identify positive hits that may be overlooked by peptide phosphorylation kinase assays. Kinase assays, such as the ones used to identify the drugs Compound C and A-592107, are biased toward the identification of ATP-competitive inhibitors that bind AMPK-α and AMP-mimetics that increase the activity of p-AMPK, respectively [4Zhou G, Myers R, Li Y, et al. Role of AMP-activated protein kinase in mechanism of metformin action J Clin Invest 2001; 108(8): 1167-74., 26Anderson SN, Cool BL, Kifle L, et al. Microarrayed compound screening (microARCS) to identify activators and inhibitors of AMP-activated protein kinase J Biomol Screen 2004; 9(2): 112-21., 27Cool B, Zinker B, Chiou W, et al. Identification and characterization of a small molecule AMPK activator that treats key components of type 2 diabetes and the metabolic syndrome Cell Metab 2006; 3(6): 403-16.]. (A-592107 was later used as a structural template for the optimized synthetic analog A-769662, but this drug was shown to be ineffective against AMPK heterotrimers containing AMPK-β2 [27Cool B, Zinker B, Chiou W, et al. Identification and characterization of a small molecule AMPK activator that treats key components of type 2 diabetes and the metabolic syndrome Cell Metab 2006; 3(6): 403-16., 28Treebak JT, Birk JB, Hansen BF, Olsen GS, Wojtaszewski JF. A-769662 activates AMPK beta1-containing complexes but induces glucose uptake through a PI3-kinase-dependent pathway in mouse skeletal muscle Am J Physiol Cell Physiol 2009; 297(4): C1041-52.]). Because purified protein phosphatases are not included in kinase activity assays, these assays are biased away from the identification of protective ADP-mimetics [4Zhou G, Myers R, Li Y, et al. Role of AMP-activated protein kinase in mechanism of metformin action J Clin Invest 2001; 108(8): 1167-74.]. Our fluorescence-based assay, on the other hand, can identify molecules that displace MANT-ADP. Orthogonal follow-up assays, including in vitro assays in the presence of phosphatases, can then determine if positive hits are allosteric inhibitors, allosteric activators, dephosphorylation inhibitors, or candidates for structure-activity relationship (SAR) optimization.
To optimize MANT-ADP’s protein-bound fluorescent signal, we tested 16 unique assay mixtures in a full-factorial screening design of experiments study (Fig. 2A). We subsequently decided to minimize our assay’s ionic strength to increase the protein-bound signal of MANT-ADP. Our full-factorial data agrees with Saiu’s and Xiao et al.’s data, which show an inverse relationship between salt concentration and nucleotide binding affinities for AMPK-γ [6Xiao B, Heath R, Saiu P, et al. Structural basis for AMP binding to mammalian AMP-activated protein kinase Nature 2007; 449(7161): 496-500., 14Saiu P. Structural and functional studies on nucleotide binding to
AMP-activated protein kinase. London: University College
London 2010. [Accessed March 2013]; Available:
http://discovery.ucl.ac.uk/64-5676/1/645676.pdf]. Published nucleotide binding studies have, in fact, been performed in low ionic strength conditions [6Xiao B, Heath R, Saiu P, et al. Structural basis for AMP binding to mammalian AMP-activated protein kinase Nature 2007; 449(7161): 496-500.]. Further characterization of positive hits from this assay should, however, be completed under more physiological conditions.
Although increasing concentrations of MANT-ADP and AMPK would have produced a larger assay window, we decided to continue with low concentrations of each so that all MANT-ADP-binding sites would be saturated by positive hits when screening libraries at 5 µM or less (Fig. 2B). Instead of maximizing the Z’-factor, we prioritized the sensitivity of the assay to small molecule detection. Despite the low signal-to-background ratio, the assay was robust and identified positive hits that dose-dependently inhibited MANT-ADP fluorescence. It is important to note that our experiments used a standard Coumarin (360 nm excitation and 460 nm emission) filter set. The greatest difference between MANT-ADP’s bound and un-bound signals, however, occurs close to 440 nm (Fig. 1B). Switching from a 460 nm to a 440 nm filter could be an alternative way to increase the assay window and/or Z’-factor without changing the biochemistry, but has the disadvantage of potentially increasing the interference of autofluorescent compounds in the near-ultraviolet range.
One substantial drawback to using MANT-ADP as a fluorescent probe is the potential for false negatives due to the high percentage of autofluorescent compounds near 460 nm [29Martin MP, Alam R, Betzi S, Ingles DJ, Zhu JY, Schonbrunn E. A novel approach to the discovery of small-molecule ligands of CDK2 ChemBioChem 2012; 13(14): 2128-36.-31Larsen RS, Zylka MJ, Scott JE. A high throughput assay to identify small molecule modulators of prostatic acid phosphatase Curr Chem Genomics 2009; 3: 42-9.]. Thus, our assay cannot distinguish between fluorescent true negatives and fluorescent false negatives that inhibit binding of MANT-ADP to AMPK. A red-shifted fluorophore, such as TNP-ADP, could reduce this problem. Substitution of 0.1 μM MANT-ADP with 0.1 μM TNP-ADP, however, failed to produce an assay window (Fig. S2). Furthermore, trinitrophenylation replaces both of the ribosyl hydroxyl groups on nucleotides [32Hiratsuka T, Uchida K. Preparation and properties of 2'(or 3')-O-(2,4,6-trinitrophenyl) adenosine 5'-triphosphate, an analog of adenosine triphosphate Biochim Biophys Acta 1973; 320(3): 635-47.]. Because these hydroxyl groups form hydrogen bonds with aspartate residues on AMPK-γ, trinitrophenylation could decrease the affinity of adenine nucleotides to AMPK-γ [9Chen L, Wang J, Zhang YY, et al. AMP-activated protein kinase undergoes nucleotide-dependent conformational changes Nat Struct Mol Biol 2012; 19(7): 716-8., 14Saiu P. Structural and functional studies on nucleotide binding to AMP-activated protein kinase. London: University College London 2010. [Accessed March 2013]; Available: http://discovery.ucl.ac.uk/64-5676/1/645676.pdf]. In their discussion of a TNP-ATP-based screening assay, Guarnieri et al. suggested using MANT-ATP as an alternative because TNP-ATP had such a weak binding affinity to their target kinase [22Guarnieri MT, Blagg BS, Zhao R. A high-throughput TNP-ATP displacement assay for screening inhibitors of ATP-binding in bacterial histidine kinases Assay Drug Dev Technol 2011; 9(2): 174-83.]. To compensate for TNP-ATP’s high KD, the authors increased their probe, protein, and library concentrations to 150, 100, and 100 μM, respectively [22Guarnieri MT, Blagg BS, Zhao R. A high-throughput TNP-ATP displacement assay for screening inhibitors of ATP-binding in bacterial histidine kinases Assay Drug Dev Technol 2011; 9(2): 174-83.]. Indeed, we had to increase TNP-ADP to 100 μM to observe a window between the probe’s protein-bound and unbound fluorescent signals (Fig. S2). A better probe may be a red-shifted Alexa Fluor-ADP analog that retains one of the ribosyl hydroxyls needed for binding. Alexa Fluor-ADP is not environment-sensitive, however, so one would need to use fluorescence polarization to detect binding of the probe to AMPK.
Finally, our library screen yielded a relatively low hit rate of 0.04% with a 60% confirmation rate. Multiple factors may have contributed to our low hit rate. First, nearly one-fourth of our library molecules produced fluorescent signals that were higher than the average DMSO control signal and, therefore, limited the screening efficiency, thereby lowering the hit rate. These molecules, which are shown on our scatter plot as having negative inhibition (Fig. 4), are most likely autofluorescent and may include false negatives that inhibit binding of MANT-ADP to AMPK. It is unlikely that these molecules could be increasing fluorescence by promoting interactions between AMPK and MANT-ADP, as the protein concentration exceeds the MANT-ADP concentration by almost 5-fold. Any MANT-ADP molecules that dissociate from one AMPK heterotrimer should bind immediately to another heterotrimer. Finally, the small molecule library we screened herein includes molecules that were pre-selected based on structural similarities to known kinase inhibitors and could, therefore, bind kinase domains [16Hutti JE, Porter MA, Cheely AW, et al. Development of a high-throughput assay for identifying inhibitors of TBK1 and IKKepsilon PLoS ONE 2012; 7(7): e41494., 17Peterson EJ, Janzen WP, Kireev D, Singleton SF. High-throughput screening for RecA inhibitors using a transcreener adenosine 5'-O-diphosphate assay Assay Drug Dev Technol 2012; 10(3): 260-8.]. If MANT-ADP binds only the nucleotide-binding sites on AMPK-γ, then the assay will be biased toward the discovery of small molecules that bind the regulatory region of AMPK and away from the discovery of promiscuous kinase modulators that bind the canonical, catalytic ATP-binding pocket found throughout the kinome. Our low hit rate, therefore, may reaffirm this bias and may be much lower than the hit rate for future screens of more diverse small molecule libraries.
CONCLUSION
This screen has identified small molecules that are capable of inhibiting MANT-ADP’s protein-bound fluorescent signal. Small molecules that dose-dependently inhibit MANT-ADP fluorescence may include AMPK inhibitors or activators. Orthogonal follow-up studies with these molecules will demonstrate if they can also displace ADP and will be performed both biochemically in vitro and then in cell lines for AMPK modulation in a cell intact physiological environment. Positive hits that bind the regulatory fragment but do not regulate AMPK may be used as structural templates for SAR optimization. Ultimately, the best way to determine if these molecules actually regulate AMPK is to test them in both cell-based assays and in vitro kinase assays before evaluating in vivo function.
CONFLICT OF INTEREST
The authors confirm that this article content has no conflicts of interest.
SUPPLEMENTARY MATERIAL
Supplementary material can be viewed at Download Supplementary Material
ACKNOWLEDGEMENTS
A National Institutes of Health Grant NS080108 to J.E.B. helped fund this work. The authors would like to thank Lawrence Forsberg for his assistance with protein purification and Ginger Smith for her assistance with library screening. The authors would also like to thank Duane Bronson of ScreenAble Solutions and Bill Janzen and other members of the CICBDD for their expertise and preparation of the small molecule library. Finally, the authors would like to thank Steve Gamblin and Uwe Schlattner for providing vectors encoding the AMPK regulatory fragment and full-length AMPK, respectively.
REFERENCES
| [1] | Carling D, Thornton C, Woods A, Sanders MJ. AMP-activated protein kinase: new regulation, new roles? Biochem J 2012; 445(1): 11-27. |
| [2] | Oakhill JS, Scott JW, Kemp BE. AMPK functions as an adenylate charge-regulated protein kinase Trends Endocrinol Metab 2012; 23(3): 125-32. |
| [3] | Kim M, Tian R. Targeting AMPK for cardiac protection: opportunities and challenges J Mol Cell Cardiol 2011; 51(4): 548-3. |
| [4] | Zhou G, Myers R, Li Y, et al. Role of AMP-activated protein kinase in mechanism of metformin action J Clin Invest 2001; 108(8): 1167-74. |
| [5] | Handa N, Takagi T, Saijo S, et al. Structural basis for compound C inhibition of the human AMP-activated protein kinase alpha2 subunit kinase domain Acta Crystallogr D Biol Crystallogr 2011; 67(Pt 5): 480-7. |
| [6] | Xiao B, Heath R, Saiu P, et al. Structural basis for AMP binding to mammalian AMP-activated protein kinase Nature 2007; 449(7161): 496-500. |
| [7] | Xiao B, Sanders MJ, Underwood E, et al. Structure of mammalian AMPK and its regulation by ADP Nature 2011; 472(7342): 230-3. |
| [8] | Oakhill JS, Chen ZP, Scott JW, et al. beta-Subunit myristoylation is the gatekeeper for initiating metabolic stress sensing by AMP-activated protein kinase (AMPK) Proc Natl Acad Sci USA 2010; 107(45): 19237-41. |
| [9] | Chen L, Wang J, Zhang YY, et al. AMP-activated protein kinase undergoes nucleotide-dependent conformational changes Nat Struct Mol Biol 2012; 19(7): 716-8. |
| [10] | Oakhill JS, Steel R, Chen ZP, et al. AMPK is a direct adenylate charge-regulated protein kinase Science 2011; 332(6036): 1433-5. |
| [11] | Sim AT, Hardie DG. The low activity of acetyl-CoA carboxylase in basal and glucagon-stimulated hepatocytes is due to phosphorylation by the AMP-activated protein kinase and not cyclic AMP-dependent protein kinase FEBS Lett 1988; 233(2): 294-8. |
| [12] | Chen ZP, Stephens TJ, Murthy S, et al. Effect of exercise intensity on skeletal muscle AMPK signaling in humans Diabetes 2003; 52(9): 2205-12. |
| [13] | Oakhill JS, Scott JW, Kemp BE. Structure and function of AMP-activated protein kinase Acta Physiol (Oxf) 2009; 196(1): 3-14. |
| [14] | Saiu P. Structural and functional studies on nucleotide binding to AMP-activated protein kinase. London: University College London 2010. [Accessed March 2013]; Available: http://discovery.ucl.ac.uk/64-5676/1/645676.pdf |
| [15] | Zhang JH, Chung TD, Oldenburg KR. A simple statistical parameter for use in evaluation and validation of high throughput screening assays J Biomol Screen 1999; 4(2): 67-73. |
| [16] | Hutti JE, Porter MA, Cheely AW, et al. Development of a high-throughput assay for identifying inhibitors of TBK1 and IKKepsilon PLoS ONE 2012; 7(7): e41494. |
| [17] | Peterson EJ, Janzen WP, Kireev D, Singleton SF. High-throughput screening for RecA inhibitors using a transcreener adenosine 5'-O-diphosphate assay Assay Drug Dev Technol 2012; 10(3): 260-8. |
| [18] | Lipinski CA, Lombardo F, Dominy BW, Feeney PJ. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings Adv Drug Deliv Rev 1997; 23(1-3): 3-25. |
| [19] | Neumann D, Woods A, Carling D, Wallimann T, Schlattner U. Mammalian AMP-activated protein kinase: functional, heterotrimeric complexes by co-expression of subunits in Escherichia coli Protein Expr Purif 2003; 30(2): 230-7. |
| [20] | Rajamohan F, Harris MS, Frisbie RK, et al. Escherichia coli expression, purification and characterization of functional full-length recombinant alpha2beta2gamma3 heterotrimeric complex of human AMP-activated protein kinase Protein Expr Purif 2010; 73(2): 189-97. |
| [21] | Riek U, Scholz R, Konarev P, et al. Structural properties of AMP-activated protein kinase: dimerization, molecular shape, and changes upon ligand binding J Biol Chem 2008; 283(26): 18331-43. |
| [22] | Guarnieri MT, Blagg BS, Zhao R. A high-throughput TNP-ATP displacement assay for screening inhibitors of ATP-binding in bacterial histidine kinases Assay Drug Dev Technol 2011; 9(2): 174-83. |
| [23] | Simpson RJ. Stabilization of proteins for storage Cold Spring Harb Protoc pdb top 2010; 2010(5) pdb top79 |
| [24] | Machrouhi F, Ouhamou N, Laderoute K, et al. The rational design of a novel potent analogue of the 5'-AMP-activated protein kinase inhibitor compound C with improved selectivity and cellular activity Bioorg Med Chem Lett 2010; 20(22): 6394-9. |
| [25] | Yu PB, Hong CC, Sachidanandan C, et al. Dorsomorphin inhibits BMP signals required for embryogenesis and iron metabolism Nat Chem Biol 2008; 4(1): 33-41. |
| [26] | Anderson SN, Cool BL, Kifle L, et al. Microarrayed compound screening (microARCS) to identify activators and inhibitors of AMP-activated protein kinase J Biomol Screen 2004; 9(2): 112-21. |
| [27] | Cool B, Zinker B, Chiou W, et al. Identification and characterization of a small molecule AMPK activator that treats key components of type 2 diabetes and the metabolic syndrome Cell Metab 2006; 3(6): 403-16. |
| [28] | Treebak JT, Birk JB, Hansen BF, Olsen GS, Wojtaszewski JF. A-769662 activates AMPK beta1-containing complexes but induces glucose uptake through a PI3-kinase-dependent pathway in mouse skeletal muscle Am J Physiol Cell Physiol 2009; 297(4): C1041-52. |
| [29] | Martin MP, Alam R, Betzi S, Ingles DJ, Zhu JY, Schonbrunn E. A novel approach to the discovery of small-molecule ligands of CDK2 ChemBioChem 2012; 13(14): 2128-36. |
| [30] | Jarecki J, Chen X, Bernardino A, et al. Diverse small-molecule modulators of SMN expression found by high-throughput compound screening: early leads towards a therapeutic for spinal muscular atrophy Hum Mol Genet 2005; 14(14): 2003-18. |
| [31] | Larsen RS, Zylka MJ, Scott JE. A high throughput assay to identify small molecule modulators of prostatic acid phosphatase Curr Chem Genomics 2009; 3: 42-9. |
| [32] | Hiratsuka T, Uchida K. Preparation and properties of 2'(or 3')-O-(2,4,6-trinitrophenyl) adenosine 5'-triphosphate, an analog of adenosine triphosphate Biochim Biophys Acta 1973; 320(3): 635-47. |